CURRENT RESEARCH

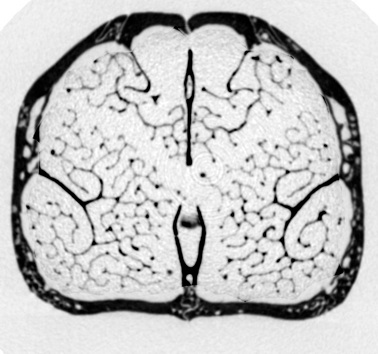
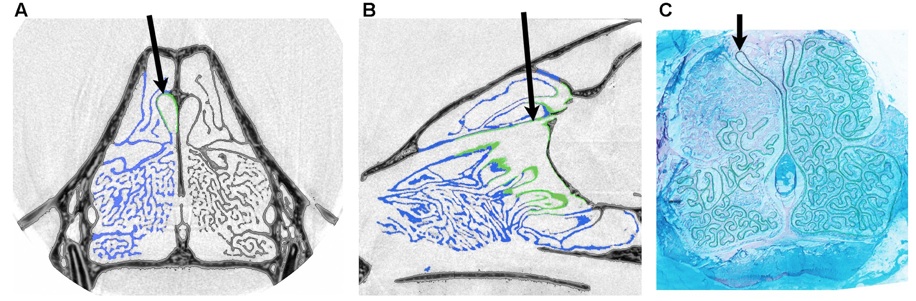
A major area of my current research involves the study of turbinate (both respiratory and olfactory) structure across the entire order Carnivora. Nearly all mammals possess complex bony scrolls within the nasal chamber, the maxilloturbinates, which function to conserve water and warm inspired air during ventilation. Olfactory turbinates are adjacent to respiratory turbinates but are covered in sensory epithelium. Despite the fact that olfactory and respiratory turbinates make up as much as a third of a mammalian skull, variations in their structure are poorly understood, largely because the fine-scale and internal location of turbinates make them difficult to visualize. Funded by NSF, we are using enhanced computer visualization of high-resolution CT scans to quantify turbinate surface area and other parameters in 30+ species across the order Carnivora (click here for a list of scanned species: CT scanned carnivora.pdf). Working with Dr. S. Soatto and his graduate students in the Computer Science Department at UCLA, we developed new software to resolve these complex structures and virtually extract them in three dimensions from the skull such that aspects of their anatomy can be readily quantified. In collaboration with engineers at Penn State, we are using computational fluid dynamics to quantify airflow in the nasal cavity. Ultimately, insights from this work will be transferable to fossil mammals, assisting us in reconstructing the evolutionary history of two fundamental features of mammalian evolution: olfaction and respiration.
Using tooth wear as a window into past community dynamics is a novel approach to the fossil record that I pioneered. I first explored the implications of unusually heavy tooth wear in Pleistocene carnivores in a 1993 Science paper on Rancho La Brea carnivores. More recently I compared dental fracture frequencies between several Pleistocene predator populations (Alaska, California, Texas, Mexico, Venezuela) and those of 35 species of extant carnivores, ranging from skunks to lions. The mean frequency of tooth fracture in all the Pleistocene species was four times that of modern species, and the trend was even apparent when comparing Pleistocene and living populations of the same species such as coyotes and wolves. This suggests that selection pressures on large carnivore in the pre-human dominated world may have been just as severe, although different in kind from what they experience at present and historically, and should alter our view of how pristine mammal communities functioned.
Seeking yet another novel approach to interpreting the lives of extinct species, I turned to predator playback experiments in Africa used to census lion and hyenas. In these experiments, the sounds of dying wildebeest are used to attract carnivores and the individuals are counted. I was struck by the similarity between the playbacks and the situation at Rancho La Brea in which numerous carnivores were lured to their death by a dying herbivore in the tar seeps. In the African experiments, large and small social species (lions, hyenas, jackals) actively moved towards the playbacks while large and small solitary species rarely did so. This suggested a strong advantage for sociality (safety in numbers) when approaching a carcass, given the potential danger for battle with similarly motivated individuals. Working with C. Carbone and others, we showed that the parallels between La Brea and Africa were remarkable, assuming that the sabertooth cat (Smilodon) was social. Instead of hyenas, lions, and jackals, it was dire wolves, Smilodon, and coyotes. It is very difficult to infer social behavior in extinct species, and our paper provides the strongest inferential evidence for sociality in a sabertooth cat thus far.
Given that coexistence among carnivores is favored by differences in diet, I continue to be intrigued by the evolution of feeding adaptations. As part of an international symposium on extreme feeding adaptations in vertebrates, my student G. Slater and I undertook a multivariate shape analysis of sabertooth “cat” skulls, including both an extinct family of nimravid “cats” as well as the family Felidae. Our paper, published in Paleobiology, revealed significant parallels in the evolution of the sabertooth form in both families, with each producing two distinct types, short and long-toothed. Moreover, within each of these, the trajectory of shape change over time appeared to be driven largely by selection for increased gape (bite size). For a separate symposium on feeding adaptations, I reviewed the wider history of iterative evolution in carnivore feeding types, highlighting the repeated, independent incarnations of dog-like, cat-like, bear-like and hyena-like forms over the past 50 million years. These iterations strongly suggest constraints on predator evolution incurred by competition.
Trade-offs in function also act as constraints on carnivore skull form. For example, adaptations that favor bite strength are opposed to those that favor jaw-closing speed. To explore this trade-off, G. Slater, E. Dumont and I applied FEA to the skulls of three wild canid species, and were able to document differences in bone strain distribution and magnitude that were engendered by skull shape. Unlike many previous FEA studies of cranial form that tend to focus on only one spectacular species such as great white sharks, ours is the first to take an explicitly comparative and evolutionary approach. Without this, it is difficult to infer the pathways taken by natural selection in producing a particular skull form.
To understand the evolution of adaptation, it is critical to have a well-resolved evolutionary tree. To this end, I am co-PI (with 2 others and 10 additional senior scientists) on an NSF funded endeavor to unravel the evolutionary history of the order Carnivora. As part of this effort, we have nearly completed a species-level evolutionary tree for the entire order based on data from multiple genes (for >280 species). Concurrently, we are assembling sets of morphological characters that are expected to be informative concerning either systematic relationships and/or functional evolution. Our goals are several but include determining the relative value of different types of characters (e.g. cranial, dental, post-cranial) in determining evolutionary relationships as well as testing a variety of hypotheses concerning functional evolution in the order. Although we are focused on the order Carnivora, our results are likely to be applicable to the study of evolution within other mammalian orders.
Molecular and morphological evolution within the Carnivora.
Evolution of feeding adaptations
Parallels between past and present predator guilds
Function and evolution of mammalian turbinates